



The role of T-cells in the pathogenesis of chronic lymphocytic leukemia has recently gained much attention due to the importance of the constant interaction between neoplastic B-cells with microenvironment substratum and T-cells. It is believed that these interactions modulate the clinical course of the disease, mainly through the regulation of the expansion, differentiation, and survival of chronic lymphocytic leukemia B-cells. Importantly, this crosstalk may also change the number, function, and memory phenotype of normal T-cells, thereby altering the amplitude and/or efficiency of adaptive immunity in chronic lymphocytic leukemia patients. The present study presents an overview on important aspects of this immunological crosstalk, particularly on the abnormalities of chronic lymphocytic leukemia B-cells and the alterations in normal T-cells, with focus on the CD4 memory T-cell compartment that could offer survival signals to chronic lymphocytic leukemia B-cell clone(s) and contribute to the establishment and progression of the disease. The authors believe that understanding the biological consequences of the interaction between normal T- and neoplastic B-cells in chronic lymphocytic leukemia may allow for improvements in the prognostic information and therapeutic approaches for this disease.
© 2014 Associação Brasileira de Hematologia, Hemoterapia e Terapia Celular. All rights reserved.
Chronic lymphocytic leukemia (CLL) is the most common mature B-cell neoplasm in Western countries. It is characterized by the appearance of monoclonal CD5+CD19+ mature B-cells in the peripheral blood, lymphoid system, and bone marrow.1 The prevalence of the disease is higher in men compared to women and the estimated incidence is two to six cases per 100,000 people annually. At the time of diagnosis, approximately 31% of the patients are younger than 64 years and the average age is 72 years.1,2
The clinical course of CLL is heterogeneous, and survival can vary from months to decades. Although most patients have an asymptomatic disease, there is a group of patients with aggressive CLL characterized by autoimmune hemolytic anemia, recurrent infections, immunodeficiency, and transformation to aggressive lymphoma, with an average life expectancy of less than three years.1,3
Several factors play an important role in the etiology of CLL, such as genetic predisposition related to the familiar history, environmental factors, and antigens/auto-antigens promoting division of precursor cells and clonal evolution.1,4 Moreover, the study of CLL genome by sequencing approaches revealed novel mutated genes, such as MYD88, NOTCH1, SF3B1, and XPO1, among others. Importantly, some of these genes may be considered as prognostic factors.5-6
Another important aspect that modulates the outcome of the disease in CLL patients is the interaction between neoplastic B-cells with microenvironment substratum and T-cells. These interactions occur in organized structures termed pseudofollicular proliferative centers (PC), which are clusters of small lymphocytes dispersed in lymph nodes and bone marrow. Interestingly, PC are not visualized in any other B-cell neoplasm, and this structure is considered a hallmark of CLL.7 Data in the literature suggest that the crosstalk between CLL B-cells, extracellular components of the microenvironment, and T-cells has an important impact on the physiopathology and evolution of the disease, mainly through regulation of CLL B-cell expansion, differentiation, and survival. Conversely, this crosstalk may also induce qualitative and quantitative changes in normal T-cells that could impact the fitness of the immune system of CLL patients.7-10
Tables 1 and 2 summarize some characteristics of neoplastic B-cells and normal T-cells, respectively, that may impact in CLL physiopathology.
Characteristics of CLL B-cells.
| Characteristics | Implications |
| Reduction in D80/CD86 expression | CLL B-cells are poor antigen presenting cells11 |
| CD200 expression | Inhibition of T-helper-1 and induction of regulatory T-cells (Tregs)12,13 |
| Expression of FASL with downregulation of FAS | Protection of CLL B-cells from FAS-mediated cell death; promotion of T-cell apoptosis9 |
| Increase of soluble FAS | Related to the progressive CLL14 |
| Secretion of soluble interleukin-2 receptor and interleukin-10 | Inhibition of T-helper-1 differentiation15,16 |
| Interleukin-6 secretion | Protection of CLL B-cells from spontaneous apoptosis; secretion of interleukin-4 by T-cells and consequent positive impact on CLL B-cell survival17,18 |
CLL: chronic lymphocytic leukemia.
Qualitative and quantitative changes in T-cells induced by CLL B-cells.
| Changes in T-cell | Comments |
| Increase in CD4 and CD8 T-cell absolute number | Due to leukocytosis9,10,19 |
| Inversion of the CD4/CD8 ratio in peripheral blood | CD4 T-cells are more sensitive to FAS mediated cell death20,21 |
| Elevated expression of FAS | Increased rates of T-cell apoptosis induced by FASL on CLL B-cells20 |
| Deficiency acquired in CD40L expression | CLL cells induce downregulation of CD40L on the T-cell surface22 |
| Abnormal profile of cytokine and/or cytokine receptor expression10 | - |
| Expansion of regulatory T-cells (Tregs) | Induced by CD200 expression in CLL B-cells12,13 |
| Reduction in T-cell receptor repertoire | T-cell oligloclonal expansion23 |
| Interleukin-4 secretion | Induced by interleukin-6 secretion of CLL B-cells17,18,24 |
| Cytoskeleton changes | Defective immune synapse25 |
| Alterations in genes involved in CD8 T-cell cytotoxicity | Inefficient CD8 T-cell cytotoxicity26,27 |
| Defective lymphocyte functionassociated antigen 1-directed T-cell motility | CLL B-cells alter the Rho GTPase signaling28,29 |
CLL: chronic lymphocytic leukemia.
In addition to these changes in the T-cell compartment, recent data suggest an accumulation of memory T-cells in CLL patients that is associated with a more aggressive course of the disease.19,21,30,31 Therefore, this study aimed to discuss the possible involvement of memory T-cells in the physiopathology and clinical course of CLL.
Memory T-cellsT-cells play a crucial role in the immune system; they are critical for combating and controlling tumors and intracellular and extracellular pathogens, acting as cytotoxic cells (cytotoxic T-lymphocytes [CTL]) or assisting other immune cells (T-helper [Th] lymphocytes). Importantly, Th lymphocytes differentiate into subsets capable of producing different cytokine patterns and, therefore, exerting diverse helper functions.8
The course of immune response can be briefly summarized by initial antigen-specific stimulation of naîve T-cells that results in activation, vigorous proliferation, and differentiation to specific effector T-cell subpopulations, which are capable of fighting pathogens and tumor cells.8
After pathogen clearance, the majority of effector T-cells die due to lack of stimulation with participation of the proapoptotic protein B-cell lymphoma 2 interacting mediator of cell death (BIM).32 In the case of chronic activation of T-cells, these may undergo activation-induced cell death (AICD), a cell death regulatory mechanism mediated by the FAS/ FASL interaction.33-34 Together, this homeostatic mechanism attempts to restore the initial ‘baseline’ immune system and is defined as the contraction phase of immune response.
However, a small fraction of antigen-specific T-cells are resistant to cell death and become long-lived memory T-cells. These cells comprise a heterogeneous group that are more sensitive to low antigen concentrations and that survive in the body to give faster and more effective antigen-specific responses, providing immediate protection in peripheral tissues and the ability to confer secondary responses in lymph nodes.34
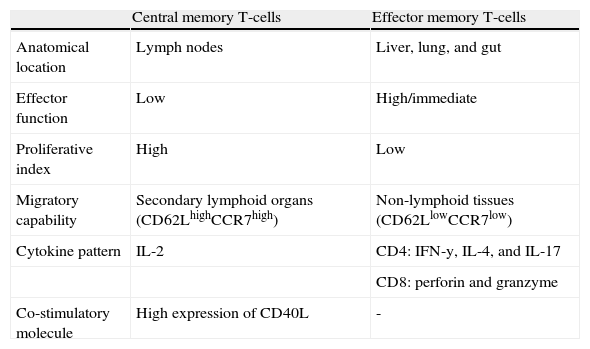
Criteria such as immediate effector function, phenotypic diversity, proliferative index, migratory capability, and anatomical location are used to define memory T-cells in human models and characterize them in central memory (TCM) and effector memory (TEM) profiles (Table 3). This classification was proposed in 1999 by Sallusto et al. and is now widely accepted in the international scientific literature.35,36
Central and effector memory T-cell characteristics.
| Central memory T-cells | Effector memory T-cells | |
| Anatomical location | Lymph nodes | Liver, lung, and gut |
| Effector function | Low | High/immediate |
| Proliferative index | High | Low |
| Migratory capability | Secondary lymphoid organs (CD62LhighCCR7high) | Non-lymphoid tissues (CD62LlowCCR7low) |
| Cytokine pattern | IL-2 | CD4: IFN-y, IL-4, and IL-17 |
| CD8: perforin and granzyme | ||
| Co-stimulatory molecule | High expression of CD40L | - |
CLL: chronic lymphocytic leukemia.
As previously mentioned, constant interaction with the microenvironment substratum and T-cells is essential for CLL B-cells in order to avoid apoptosis and acquire favorable growing conditions. As a result of this crosstalk, some changes in T-cells are well documented, but a recent observation that CLL is able to interfere in naîve and memory T-cell status deserves more discussion.19,21,30,31
It has been demonstrated that CLL patients have a significant skewing only in the CD4 T-cell compartment towards TCM and TEM cells.37 Interestingly, this altered T-cell profile was associated with a more aggressive course of the disease, as shown by the positive association with unmutated human immunoglobulin heavy chain variable genes (IgVH), advanced regent admission index clinical stage, and shorter treatment-free survival. Conversely, there was no correlation between the increase in CD4 memory T-cells and CD38 expression or genomic aberrations in CLL patients.19
Another study demonstrated that a CLL mouse model had decreased naîve T-cells with concomitant increase in antigenexperienced memory T-cells, mainly the subtypes that have the ability to migrate to lymph nodes.30 As the background of the mouse model used was the TCL1, which represents an aggressive CLL, it is possible that the increased numbers of memory T-cells are related to the aggressiveness of CLL, both in humans and mice.
An interesting question is how the different types of memory T-cells could influence CLL physiopathology. It has been postulated that chronic antigenic stimulation through the B-cell receptor (BCR) is required for the neoplastic clone to survive and grow.38 In this context, CD4 TCM cells have the capability to migrate to the lymph nodes due to their expression of CXCR5 (a chemokine CXCL13-receptor produced by B-cells in lymph node follicles), and could interact with CLL B-cells, providing co-stimulatory signals, such as CD40L and cytokines.36 This crosstalk could contribute to a stronger BCR signaling, greater survival, expansion, and evolution of CLL.
In addition, according to the literature on memory T-cell generation, a low but sufficient activation of naîve T-cells is crucial to the differentiation of TCM cells.39 Associating this evidence with the fact that CLL B-cells are poor as antigen presenting cells,11 the interaction between CLL B-cells and naîve T-cells could result in a weak stimulation leading to the generation and/or accumulation of CD4 TCM cells.
Regarding TEM cells, cytokine production of interleukin-4 (IL-4) and interferon-gamma (IFN-γ) could also participate in CLL progression, mainly through the up-regulation of BCL-2 and protection of CLL cells from apoptosis.40,41 It was demonstrated that, in in vitro co-culture assays, the apoptosis rate of CLL B-cells in the presence of CD4 TEM cells was lower than in other conditions, such as in the presence of either naîve CD4 T-cells or peripheral blood mononuclear cells. Moreover, the protection conferred by CD4 TEM cells seems to depend on IL-4.19
Considered together, the existence of an intrinsic feedback mechanism involved in the physiopathology of the CLL is supported, in which CLL B-cells could induce the generation and/or accumulation of CD4 memory T-cells, which, in turn, would help them achieve better survival and expansion (Figure 1). Indeed, it was demonstrated that CD4 memory T-cell generation is impaired in mice that are deficient in B-cells.37,42

At the lymph node site, neoplastic B-cells interact with naîve and activated CD4 T-cells, resulting in generation/ accumulation of CD4 memory T-cells in peripheral blood. Through the lymph node homing receptors, chemokine receptors and co-stimulatory CD4 central memory T-cells could offer survival signals to chronic lymphocytic leukemia B-cells at lymph node sites. In addition, cytokines produced by CD4 effector memory T-cells, such as interleukin-4 and interferon- gamma, could also participate in this survival process.
Unpublished data from our group supports the aforementioned information. Briefly, we analyzed the peripheral blood T-cells of 21 CLL patients, and evaluated whether the distribution of naîve and memory T-cells was related to the ZAP-70 expression, a well-established prognosis factor in CLL.43 Interestingly, this analysis demonstrated that ZAP-70 positive patients with neoplastic B-cells presented increased frequency and absolute numbers of CD4 TCM cells compared to the ZAP-70 negative patients and 17 age-matched healthy individuals (Correia RP, unpublished data). Although other clinical and laboratorial prognostic markers were not analyzed at this point, the results are in accordance with the literature that shows alterations in the memory CD4 T-cell subpopulations. Importantly, it is suggested that this alteration is restricted to the T-helper compartment, since no significant differences were observed among memory CD8 T-cells.
To the authors’ knowledge, there is no study that evidences associations between genetic factors and T-cell skewing towards memory status. Because the authors believe that this information is critical for a better evaluation of the prognostic value of analyzing the memory T-cell compartment, as well as for the understanding of the involvement of CD4 memory T-cells in the pathophysiology of CLL, studies with this focus are underway in this laboratory.
ConclusionsAbnormalities in T-cell subsets may be associated with the progression of CLL. Particularly, the increased CD4 memory T-cells could help CLL B-cells to achieve better fitness, i.e., more survival and proliferation signals. Therefore, understanding and unraveling this mechanism could improve the prognostic information and therapeutic approaches in CLL.
Conflicts of interestThe authors declare no conflicts of interest.



