
The Kidd blood group system has three antigens, Jka, Jkb and Jk3, found on red blood cells and on endothelial cells of the inner lining of blood vessels in the renal medulla. These are known as urea transporter B (UT-B). Researchers have found that individuals carrying the Jk(a−b−) or Jk-null (UT-B null) phenotypes have a lower urine-concentrating capability and risk of severe renal impairment. This study evaluated the distribution of the Kidd phenotypes in patients with chronic kidney disease and a possible association of Kidd antigens with the development of renal disease.
MethodsJka and Jkb antigens were phenotyped using the gel column agglutination test (ID-cards Bio-RAD) in 197 patients with chronic kidney disease and 444 blood donors, as the control group. The phenotype and antigen frequencies between patients and controls were evaluated using the Chi-square method with Yates correction and logistic regression after adjustments for gender and age.
ResultsNo differences were observed between the Kidd phenotypes frequency distribution between patients with chronic kidney disease and blood donors [Jk(a−b+)=22.3% and 27.2%; Jk(a+b−)=30.5% and 24.3%; Jk(a+b+)=47.25% and 48.4%, respectively].
ConclusionThe distribution of Kidd phenotypes found in the studied population is expected for Caucasians; Jka and Jkb antigens and phenotypes were not found to be related to susceptibility for chronic kidney disease.
The red blood cell (RBC) membrane contains many anchored surface proteins and proteins that cross the lipid bilayer carrying different blood group antigens. Currently, 36 systems1 of RBC groups have been described according to the International Society of Blood Transfusion (ISBT) (http://www.isbtweb.org). Among them, the Kidd blood group system (JK; ISBT 009) has been recognized as clinically important since its identification in 1951.2
Antigens of the Kidd blood group system are expressed on type 3 glycoproteins, also known as the urea transporter B (UT-B). This protein contains 389 amino acids and passes ten times through the lipid bilayer with both the N terminus and C terminus being intracellular. Three antigens have been found (Jka, Jkb and Jk3) on the neighboring fourth extracellular looping and three phenotypes, Jk(a+b−), Jk(a−b+), and Jk(a+b+), are common among different populations. The Jk(a−b−) phenotype is rare in most populations. It was first found in a Filipina of Spanish and Chinese ancestry with the antibodies usually being detected after transfusions or pregnancies. After immunization, anti-Jk3 can be found in patients with the Jk(a−b−) recessive phenotype, causing acute and delayed hemolytic transfusion reactions.3,4
The Jk glycoprotein is coded by the Solute carrier family 14, member A1 (SLC14A1) gene, a member of the urea-transporter gene family, located on chromosome 18q12-q21 and organized into 11 exons. The two major codominant alleles of the gene, JK*A and JK*B, have similar frequencies in Caucasian populations (0.51 and 0.49, respectively) and define the Jka and Jkb antigens, respectively. The Jka/Jkb polymorphism is defined by 838 A>G in exon 9 (Asp280Asn substitution), the other two single nucleotide polymorphisms (SNPs) cause no changes to the amino acid sequence.5
The Kidd antigens or UT-B are expressed on RBCs and the endothelium of the vasa recta and epithelial surfaces of the renal inner medulla, as well as in non-renal tissues and endothelial cells.6–8 The Kidd protein is a major transporter of urea across the RBC membrane and this rapid process helps maintain the osmotic stability of the cell. Null phenotype individuals lack this transporter. Within the Kidney, the urea transporter enables the renal medulla to maintain a high concentration of both urea and urine, as well as conserve water. Individuals with the Jk(a−b−) phenotype have lower urine-concentrating ability.9–11 In UT-B-null mice, long-term UTB deficiency was associated to severe renal dysfunction and structural damage.12 UT-B isoforms are also important in several cellular functions, including urea nitrogen salvage in the colon, nitric oxide pathway modulation in the hippocampus, and normalization of the cardiac conduction system.13
Chronic kidney disease (CKD) is a major public health problem, defined as abnormalities of kidney structure and/or function, present for at least three months with implications on health.14 The adverse outcomes of the disease include loss of kidney function, cardiovascular disease and premature death. Besides environmental factors, genetic abnormalities are also involved15 including variations in the MYH9 (encoding non-muscle myosin IIA heavy chain),16APOL1 (apolipoprotein L1),17–19NPHS1 (nephrin),20 and SHROOM3 (shroom family member 3)21 genes.
The relationship between the Kidd antigens and chronic kidney disease remains unknown. Therefore, the aim of this study was to investigate the distribution of Kidd phenotypes in patients with the chronic kidney disease and a possible association of Kidd antigens with the development of renal disease.
MethodsThe ethical and methodological aspects of this study were approved by the Research Ethics Committee on Human Beings from the Maringa State University (COPEP-UEM # 1.141.385/2014, CAAE 43117115.0.0000.0104, according to the Resolution of the Brazilian Council on Health-CNS 466/12.
SubjectsThis retrospective case–control study enrolled 197 unrelated patients with chronic kidney disease (CKD group) and 444 unrelated blood donors as a control group, living in the same geographical area as the patients. The individuals were attended and immunophenotyped between 2013 and 2015 at the Regional Blood Bank of Pato Branco, southwest region of Parana (located in the southern region of Brazil at 26°13′46″–09″S and 52°40′16″–09″W).
Serologic testsRed blood cell phenotyping for Kidd blood group systems was performed on a gel card (ID-Perfil II – k-Kpa-Kpb-Jka-Jkb-ctl) using monoclonal antibodies according to the manufacturer's instructions (Diamed ID-Cards, DiaMed® AG, Switzerland). RBCs were suspended in Bromelin solution (BioRad ID-Diluent 1) at a final concentration of 1:21 or 5%.
Statistical analysisThe antigen and phenotype frequencies were estimated and the data was tested for their fit to the Hardy–Weinberg equilibrium22 by calculating the expected frequencies of the genotypes and comparing them with the observed values. The Student's t-test was used to compare differences between groups. Statistical comparisons between these groups were performed and the estimated risk of developing CKD in individuals who have genetic polymorphisms was calculated by determining the Odds Ratio (OD) with a 95% of confidence interval (CI) adjusted for gender and age. Association tests were carried out to identify the codominant, dominant, recessive, overdominant and log-additive genetic inheritance models. p-Values≤0.05 by the Chi-square test with Yates correction and logistic regression were considered statistically significant. All statistical analyses were performed using the software OpenEpi program Version 2.3.1 (http://www.openepi.com) and SNPStats23 (http://bioinfo.iconcologia.net/index.php).
ResultsThe Kidd phenotype frequency distribution in the studied populations was in Hardy–Weinberg equilibrium (p-value=0.48: CKD and p-value=0.51: controls).
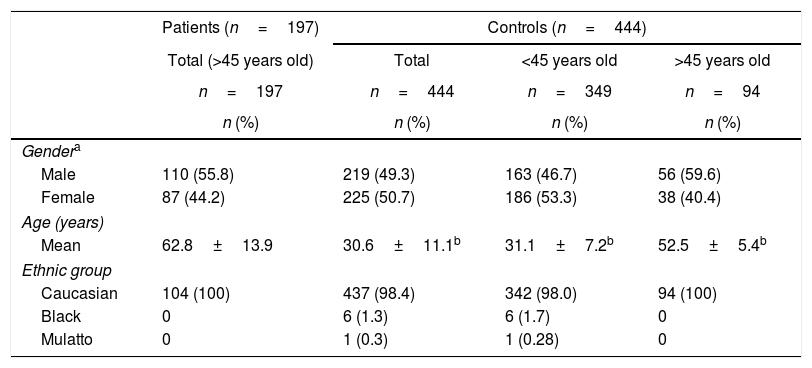
The characteristics of patient and control subjects are described in Table 1. The CKD patients were between 45 and 90 years old (62.8±13.9) and from both genders (male: 55.8%; female: 44.2%). Regarding ethnicity, all patients declared themselves Caucasians. The control group was formed by 444 individuals between 18 and 64 years old (30.6±11.1; p-value<0.0001) from both genders (1:1) with 98.4% declaring themselves Caucasians. Because CKD was observed only in individuals>45 years old, the control group was subdivided (n=94) for analyses, considering the minimum age reported by patients. In this group, the mean age was 52.54±5.36 years (p-value<0.0001), and 58.9% were males and 41.1% females with all of them being self-declared Caucasians.
Characteristics of the CKD patient and control individuals.
| Patients (n=197) | Controls (n=444) | |||
|---|---|---|---|---|
| Total (>45 years old) | Total | <45 years old | >45 years old | |
| n=197 | n=444 | n=349 | n=94 | |
| n (%) | n (%) | n (%) | n (%) | |
| Gendera | ||||
| Male | 110 (55.8) | 219 (49.3) | 163 (46.7) | 56 (59.6) |
| Female | 87 (44.2) | 225 (50.7) | 186 (53.3) | 38 (40.4) |
| Age (years) | ||||
| Mean | 62.8±13.9 | 30.6±11.1b | 31.1±7.2b | 52.5±5.4b |
| Ethnic group | ||||
| Caucasian | 104 (100) | 437 (98.4) | 342 (98.0) | 94 (100) |
| Black | 0 | 6 (1.3) | 6 (1.7) | 0 |
| Mulatto | 0 | 1 (0.3) | 1 (0.28) | 0 |
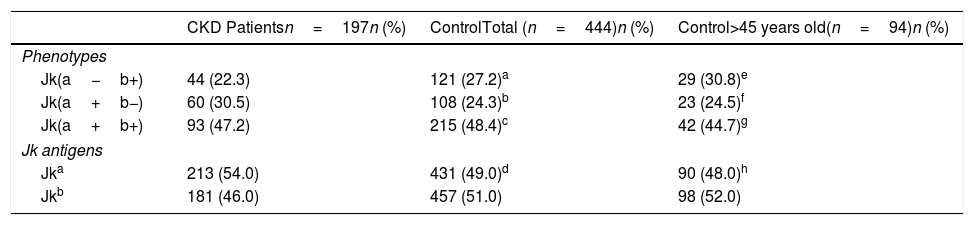
The distributions of antigen and phenotype frequencies for the Kidd blood group systems in CKD patients and controls (total group and group >45 years old) are shown in Table 2. The phenotype frequencies for the Kidd blood group system in CKD patients were 22.3% for Jk(a−b+), 30.5% for Jk(a+b−) and 47.2% for Jk(a+b+). For controls (total and >45 years old, respectively) the phenotype frequencies were 27.2% and 30.8% for Jk(a−b+), 24.3% and 24.5% for Jk(a+b−), and 48.4% and 44.7% for Jk(a+b+). The null phenotype was not observed in any of the groups.
Distribution of Jk antigen and phenotype frequencies in CKD patients and controls (total and >45 years old).
| CKD Patientsn=197n (%) | ControlTotal (n=444)n (%) | Control>45 years old(n=94)n (%) | |
|---|---|---|---|
| Phenotypes | |||
| Jk(a−b+) | 44 (22.3) | 121 (27.2)a | 29 (30.8)e |
| Jk(a+b−) | 60 (30.5) | 108 (24.3)b | 23 (24.5)f |
| Jk(a+b+) | 93 (47.2) | 215 (48.4)c | 42 (44.7)g |
| Jk antigens | |||
| Jka | 213 (54.0) | 431 (49.0)d | 90 (48.0)h |
| Jkb | 181 (46.0) | 457 (51.0) | 98 (52.0) |
ap-Value=0.32; bp-Value=0.55; cp-Value=1; dp-Value=0.16 (CKD patients vs. controls−total of samples).
ep-Value=0.55; fp-Value=0.18; gp-Value=1; hp-Value=0.16 (CKD patients vs. controls >45 years old).
Differences in distributions of the Jka and Jkb antigen and phenotype frequencies were not observed when case and control groups were compared according to inheritance models or after logistic regression stratified by age.
DiscussionIn this association study, patients with CKD on hemodialysis were analyzed in order to investigate the distribution of JK phenotypes in patients with CKD and a possible influence of Jka and Jkb antigens and phenotypes in the development of renal disease. However, differences between Jka or Jkb antigen and phenotype frequencies were not observed between patients and controls.
The patients’ ages ranged from 45 to 95 years old (55.8±13.9) and the predominant gender and ethnic group were male (62%) and Caucasian (100%). When the age range of CKD patients was compared with that of the control group, significant differences were found. This was because the control group was formed of blood donors from the Pato Branco Blood Bank with ages ranging between 18 and 64 years (BRASIL, ordinance 2712 of November 12, 2013). Therefore, the great majority of control individuals (78.8%) were between the ages of 18 and 44, and none of the CKD patients was under 45 years of age. On the other hand, 44.7% of the patients were between the ages of 65 and 90, but none of the control group were this old. In order to circumvent these differences in age distribution, a control subgroup was defined with only individuals over 45 years of age. However, differences between Jka or Jkb antigen and phenotype frequencies were not observed between the two control groups (total of individuals and >45 years old).
Non-matching between patients and controls with respect to age may be a weakness of this study. This bias may occur because in complex diseases many genetic and environment factors contribute to the outcome; individuals from the control group with an age of under 45 are possible candidates for developing renal dysfunction in the future. On the other hand, in case–control studies, ideally there should be at least one control per case,24 and the control group of individuals older than 45 years had fewer patients than the CKD Group. However, the Jka or Jkb antigen and phenotype frequencies in both control groups of this study were similar to those frequencies found in another Brazilian population,25 thereby validating the results in this study.
The urea transporter in the kidney enables its medulla to maintain a high concentration of both urea and urine, as well as conserve water. Individuals with the Jk(a−b−) phenotype had lower urine-concentrating ability. In animal models, long-term UTB deficiency was associated with severe renal dysfunction and structural damage. On the other hand, genetic variations of the SLC14A1 gene were associated with bladder cancer and, some genotypes were associated with higher morbidity,26–29 and rejection after renal transplant.30 However, we did not find any null alleles in CKD patients or controls that could be identified as a risk factor for the disease.
In this study, more than 50% of the CKD patients were homozygous carrying either Jk(a+b−) or Jk(a−b+) phenotypes, in agreement with the results reported in the literature for Caucasians.25 Furthermore, no differences were observed between patients and controls regarding the distribution of Jka or Jkb antigen and phenotype frequencies. Thus, Kidd phenotypes were not associated to CKD.
ConclusionThe distribution of Kidd phenotypes found in the studied population was as expected for Caucasians and therefore, Jka and Jkb antigens and phenotypes were not found to be associated to the development of CKD.
Conflicts of interestThe authors declare no conflicts of interest.
The authors appreciate the participation and cooperation of all volunteers (patients and controls), as well as of the Regional Blood Bank of Pato Branco and HEMEPAR-Centro de Hematologia e Hemoterapia do Paraná, and Immunogenetics Laboratory (LIG-UEM).



