







Sickle cell disease (SCD) is a monogenic disease and it is estimated that 300,000 infants are born annually with it. Most treatments available are only palliative, whereas the allogeneic hematopoietic stem cell transplantation offers the only potential cure for SCD.
ObjectiveGeneration of human autologous cells, when coupled with induced pluripotent stem cell (iPSC) technology, is a promising approach for developing study models. In this study, we provide a simple and efficient model for generating hematopoietic cells using iPSCs derived from a sickle cell anemia patient and an inexpensive in-house-prepared medium.
MethodThis study used iPSCs previously generated from peripheral blood mononuclear cells (PBMCs) from a patient with sickle cell anemia (iPSC_scd). Hematopoietic and erythroid differentiation was performed in two steps. Firstly, with the induction of hematopoietic differentiation through embryoid body formation, we evaluated the efficiency of two serum-free media; and secondly, the induction of hematopoietic stem/progenitor cells to erythroid progenitor cells was performed.
ResultsThe patient-specific cell line generated CD34+/CD45+ and CD45+/CD43+ hematopoietic stem/progenitor cells and erythroid progenitors, comprising CD36+, CD71+ and CD235a+ populations, as well as the formation of hematopoietic colonies, including erythroid colonies, in culture in a semi-solid medium.
ConclusionIn conjunction, our results described a simple serum-free platform to differentiate human the iPSCs into hematopoietic progenitor cells. This platform is an emerging application of iPSCs in vitro disease modeling, which can significantly improve the search for new pharmacological drugs for sickle cell disease.
Induced pluripotent stem cells (iPSCs) are cell lines generated from somatic cell reprogramming and are phenotypically similar to embryonic stem cells (ESCs). As a cell line that can be obtained in vitro from somatic cells, iPSCs are a possible alternative to circumvent the ethical issues associated with ESCs.1 Additionally, iPSCs are able to quickly self-renew and have a great potential for differentiation and are therefore considered a ready-to-use cell source.
The generation of human autologous cells, when coupled with iPSC technology, is a promising approach for developing disease models. For example, Liu et al. showed that the hematopoietic differentiation of iPSCs can be applied for the attainment of hematopoietic precursors and progenitors.2 However, other approaches are being performed to determine the pathways that are involved in the determination of definitive hematopoiesis. These investigations could provide the generation of mature hematopoietic cells and long-term hematopoietic stem cells with multiline potential.3,4
Furthermore, the establishment of efficient hematopoietic and erythroid differentiation strategies from iPSCs, make it possible to have cells available for studies of various diseases. According to various groups, iPSC lines have been generated for blood disorders, such as hemophilia and sickle cell disease.5–11 These cell lines are tools for the establishment of protocols for genome editing techniques, drug screening or models to better understand the interaction between hematopoiesis and disease development.
Recently, our group has established a protocol for the reprogramming in iPSCs of the peripheral blood mononuclear cells from sickle cell anemia patients, using integration-free vectors.10 We designed this study to evaluate the feasibility of using a simple serum-free method to generate hematopoietic cells from iPSCs. In conjunction, our results demonstrate an efficient method to produce CD34+/CD45+ cells. Thus, this paper offers a valuable tool for studying the aspects of hematopoietic differentiation of sickle cell disease.
Material and methodsEthical aspects and iPSC lineFor this study, iPSCs generated from peripheral blood mononuclear cells from a patient with sickle cell anemia (iPSC_scd)10 was used. The commercial Embryonic Stem Cell line, WAe001-A, named H1 (WiCell Research Institute) provided a positive control for a pluripotent phenotype in the experiments. The use of the human cell line in this study was approved by the Research Ethics Committee of the Clinical Hospital of Ribeirão Preto (CAAE: 19350213.0.0000.5403 and CAAE: 61387516.9.0000.5440).
Hematopoietic and erythroid differentiationHematopoietic and erythroid differentiation was performed in two steps. First, the induction of hematopoietic differentiation through embryoid body formation was done, followed by the induction of hematopoietic stem/progenitor cells to erythroid progenitor cells. Embryoid body formation (EBF) was performed through centrifugation and conducted using two hematopoietic differentiation induction media without fetal bovine serum (STEMdiff APEL 2) (StemCell Technologies) and the in-house serum-free medium (SFM) (50% Iscove’s Modified Dulbecco’s Medium (IMDM) (Gibco), 50% Ham’s F12 (Gibco), 1% Chemically Defined Lipid Concentrate (Gibco), 1% ITS-X (Gibco), 2 mM GlutaMAX (Gibco), 450 μM 1-thioglycerol (Sigma-Aldrich), 50 μg/mL ascorbic acid (Sigma-Aldrich) and 0.5% human albumin (CSL Behring), supplemented with 0.05% polyvinyl alcohol (PVA) (Sigma-Aldrich).12,13
On Day 0 (D0), iPSC colonies were enzymatically dissociated and resuspended in the differentiation media, supplemented with 10 ng/mL BMP4 (PeproTech), 10 ng/mL bFGF (R&D) and 10 μM Y-27632 dihydrochloride. Subsequently, approximately 2–4 × 103 cells/well were plated on 96-well clear round bottom ultra-low attachment microplates (Corning®), at a final volume of 50 µL. The plates were centrifuged for EBF at 300 × g for 5 min and maintained in an incubator (37 °C, 5% CO2 and controlled humidity). On the following days, differentiation media was changed with cytokine composition alteration: D2: 10 ng/mL BMP4, 10 ng/mL bFGF, 20 ng/mL VEGF and 100 ng/mL SCF; D5 and D8: 10 ng/mL BMP4, 10 ng/mL bFGF, 10 ng/mL VEGF, 50 ng/mL SCF. Finally, on D11 it contained: 10 ng/mL BMP4, 10 ng/mL bFGF, 10 ng/mL VEGF, 50 ng/mL SCF and 20 ng/mL TPO. All the cytokines used at these times were bought from PeproTech.
On D8, cells derived from the hematopoietic differentiation process were collected for clonogenic potential evaluation, for which 104 cells/mL were inoculated in semi-solid MethoCult® H4435 (STEMCELL Technologies). After 14 days, the morphology of colonies obtained was registered by microphotography.
To initiate the erythroid maturation process, cells were collected after 8–14 days of hematopoietic differentiation. The quantity of 1 × 105 cells/mL were plated on a 24-well plate pre-coated with gelatin (STEMCELL Technologies) and cultivated with in-house SFM without PVA and supplemented with 500 μg/mL human holo-transferrin (Sigma-Aldrich), 10 ng/mL IL-3, 3 U/mL EPO and 50 ng/mL SCF (PeproTech). Media change was performed each 2–3 days and cells were collected for immunophenotypic profile determination after 10–15 days of maturation.
Immunophenotypic profile determination by flow cytometryDuring hematopoietic differentiation and erythroid maturation, cells were collected for immunophenotyping. At least 105 cells were incubated with antibodies (1:20) for 15 min and then washed with 2 mL of phosphate-buffered saline (PBS) 1×. The cell suspension was centrifuged for 3 min at 330 × g. The pellet obtained was resuspended at 200 µL and then acquired (10,000 events) on flow cytometry FACSCalibur or FACSAria (Becton Dickinson). The following antibodies (BD Bioscience) were used: anti-CD34, anti-CD36, anti-CD43, anti-CD45 anti-CD71, anti-CD235a, anti-NANOG, anti-SOX-2 and anti-OCT3/4. Data obtained were analyzed using the softwares FACSDiva and CELLQuest™ (Becton Dickinson).
ResultsCharacterization of iPSCs derived from CD34+ peripheral blood mononuclear cellsThe in vitro blood cell differentiation process can be divided into key stages in which specific signals are provided for mesodermal, hemangioblast/hemogenic-endothelium and hematopoietic stem/progenitor cell specification and further maturation (Figure 1A). In this study, we used an iPSC cell line with the mutation for sickle cell anemia (iPSC_scd), previously described.10 The iPSC_scd was cultured on the geltrex matrix in the mTeSR1 medium for more than 34 passages, maintaining its morphology in an undifferentiated state (Figure 1B), similar to the H1 cell line. The pluripotent stem cells were characterized for the presence of pluripotency markers and hematopoietic markers. They presented a high percentage of cells positive for pluripotency markers OCT3/4, NANOG and SOX2 and low expression of CD43, CD34 and CD45 hematopoietic markers (Figure 1B).
 Schematic representation of hematopoietic differentiation and erythroid maturation method. (B) Morphology of pluripotent stem cell lines H1 and iPSC_scd (40× magnification) and immunophenotypic characterization for the presence of pluripotency markers, OCT 3/4, NANOG and SOX 2 and hematopoietic markers, CD43, CD34 and CD45. Bars represent the mean ± standard deviation (iPSC_scd CD43, CD34, and CD45: N = 3).")
Hematopoietic differentiation of iPSC. (A) Schematic representation of hematopoietic differentiation and erythroid maturation method. (B) Morphology of pluripotent stem cell lines H1 and iPSC_scd (40× magnification) and immunophenotypic characterization for the presence of pluripotency markers, OCT 3/4, NANOG and SOX 2 and hematopoietic markers, CD43, CD34 and CD45. Bars represent the mean ± standard deviation (iPSC_scd CD43, CD34, and CD45: N = 3).
There are different hematopoietic differentiation protocols for iPSCs, however, all of them demonstrate low efficiency. For in vitro differentiation of iPSCs into hematopoietic cells, we evaluated a protocol based on a feeder-free and serum-free culture condition. In the first stage, spin embryoid bodies (EBs) are formed in low-attachment wells and the EBs grow and differentiate in response to exogenously added cytokines. We evaluated the formation of EBs using two media, STEMdiff APEL 2 and SFM (Figure 2). The STEMdiff APEL 2 greatly improves the initial formation of EBs. Cells cultured with the SFM medium did not form embryoid bodies 24 h after the plates were centrifuged. The formation of less uniform embryoid bodies occurred in some of the wells and only after 48 h of centrifugation (Figure 2A). The round-shaped hematopoietic stem/progenitor cells were possible to be detected in the EBs cultured from iPSC_scd, especially with SFM from day 8 to day 14, when they began to detach from the plate (Figure 2B).
 Morphology of spin embryoid bodies generated in STEMdiff APEL 2 and SFM media (40× magnification). (B) Round-shaped hematopoietic cells detected in the EBs cultured from iPSC_scd with SFM at day 8 and 14 (100× magnification).")
During the EBF, cells were collected on different days to evaluate the presence of hematopoietic markers by flow cytometry (Figure 3). In both media, it was possible to detect CD34+ cells, however, the SFM presented a better performance in the formation of CD34+ cells; 53.99 ± 24.49% (N = 4) and 38.65% (N = 1), for SFM and STEMdiff APEL 2, respectively, on day 8. Subsequently, we analyzed if these cells could produce high yields of hematopoietic cells CD45+ and CD43+. Flow cytometry analysis showed that only iPSC_scd cultured with SFM presented CD45+ cells, at about 20.44 ± 35.40% (N = 3) on D8 and 42.67 ± 37.21% (N = 3) on D14 (Figure 3) and on day 14 there was the highest population of CD34+/CD45+ cells, at about 36.4 ± 31.66% (N = 3) (Figure 4).
.")
Immunophenotypic characterization for CD34, CD45, and CD43 markers. Percentage of CD34+, CD45+ and CD43+ cells during hematopoietic differentiation cultured with SFM and STEMdiff APEL 2 media. Bars represent the mean ± standard deviation (CD34+ SFM: D0 and D14, N = 3; D8, N = 4; CD34+ STEMdiff APEL 2: D0, N = 2; CD45+ SFM: N = 3; CD43+ SFM: D0 and D14, N = 3; D8, N = 4; CD43+ STEMdiff APEL 2: D0, N = 2).
.")
Immunophenotypic characterization for double-positive markers. Percentage of the double-positive populations of CD34+/CD45+ and CD43+/CD45+ cells during hematopoietic differentiation with SFM. Bars represent the mean ± standard deviation (CD34+/CD45+: D0 and D14, N = 3; D8, N = 4; CD43+/CD45+: D14, N = 3).
Moreover, flow cytometry analysis showed that only iPSC_scd cultured with SFM presented a significant population of CD43+/CD45+ cells. On day 14, the population of CD43+/CD45+ cells at 42.28 ± 36.88% (N = 3) were detected, indicating that the emergence of CD45+ cells at 20.44 ± 35.40% (N = 3) and CD43+ cells at 36.61 ± 38.25% (N = 4), detected on day 8, were followed by the emergence of the double-positive population (Figure 4).
In addition to cell surface marker expression, we functionally assayed for progenitors, using the colony-forming unit (CFU) assay for the iPSC_scd cultured with the two media (Figure 5, Figure S1). Further characterization of the cells revealed CFU-granulocytes, CFU-macrophages and erythroid colonies after 14 days in a semi-solid medium (Figure 5A). Even though we were not able to detect CD45+ cells in the iPSC_scd cultured with the STEMdiff APEL 2 medium, these cells were able to generate colonies in the CFU assay (Figure 5B).
 SFM and collected from embryoid bodies on day 8 of differentiation, (40× and 100× magnification); and in (B) STEMdiff APEL 2 medium collected from embryoid bodies on day 8 of hematopoietic differentiation (40× magnification).")
Colony formation assay of differentiated cells. iPSC_scd cells previously cultured in (A) SFM and collected from embryoid bodies on day 8 of differentiation, (40× and 100× magnification); and in (B) STEMdiff APEL 2 medium collected from embryoid bodies on day 8 of hematopoietic differentiation (40× magnification).
After the induction of differentiation by the formation of embryoid bodies on low-adhesion dishes, cells from the embryoid bodies were cultured in a maturation medium on gelatin-treated plates. The culture of hematopoietic stem/progenitor cells in the maturation medium supplemented with cytokines for erythroid differentiation promoted the maturation of these cells in the cells of erythroid lineage. Many semi-adherent rounded cells with hematopoietic stem/progenitor-like morphology were observed in the iPSC_scd cells previously cultured in the SFM (Figure 6 A–B) and in the STEMdiff medium (Figure 6 C–D).
 and in STEMdiff APEL 2 medium (C–D) was observed (A and C, 200× magnification; B and D, 100× magnification).")
Culture of iPSC_scd cells in the maturation medium at various points of time. Presence of many semi-adhered rounded cells during maturation of cells previously cultured in SFM (A–B) and in STEMdiff APEL 2 medium (C–D) was observed (A and C, 200× magnification; B and D, 100× magnification).
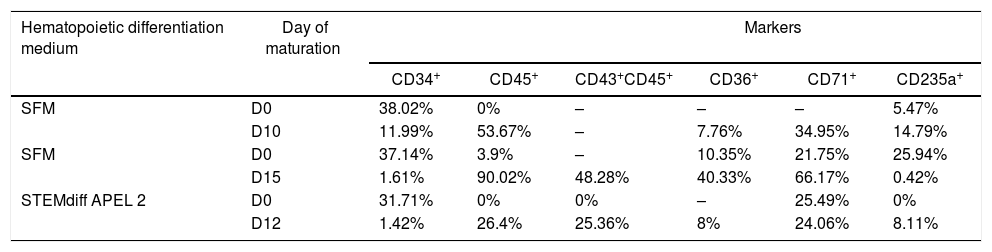
These cells were cultured for 10–15 days in a maturation medium and were characterized by flow cytometry. The analysis showed the percentage of positive cells for hematopoietic and erythroid markers CD34+, CD45+, CD43+/CD45+, CD36+, CD71+ and CD235a+ from the previously cultured cells in the SFM and in the STEMdiff APEL 2 medium (Table 1). The decrease in the CD34+ population and the increase in the presence of the other markers characterize the hematopoietic stem/progenitor cell transition and the emergence of erythroid progenitor cells.
Immunophenotypic characterization of iPSC_scd after erythroid maturation.
| Hematopoietic differentiation medium | Day of maturation | Markers | |||||
|---|---|---|---|---|---|---|---|
| CD34+ | CD45+ | CD43+CD45+ | CD36+ | CD71+ | CD235a+ | ||
| SFM | D0 | 38.02% | 0% | – | – | – | 5.47% |
| D10 | 11.99% | 53.67% | – | 7.76% | 34.95% | 14.79% | |
| SFM | D0 | 37.14% | 3.9% | – | 10.35% | 21.75% | 25.94% |
| D15 | 1.61% | 90.02% | 48.28% | 40.33% | 66.17% | 0.42% | |
| STEMdiff APEL 2 | D0 | 31.71% | 0% | 0% | – | 25.49% | 0% |
| D12 | 1.42% | 26.4% | 25.36% | 8% | 24.06% | 8.11% | |
In this study, we demonstrated that PBMC-derived iPSCs from a patient with sickle cell anemia can be differentiated in vitro into hematopoietic stem/progenitor cells and erythroid progenitors, using two serum-free media, the STEMdiff APEL 2 and the SFM. The medium STEMdiff APEL 2 (STEMCELL Technologies) is a fully defined, serum-free and animal component-free medium for the differentiation of ESCs and iPSCs. It is based on the APEL formulation published by Ng et al.13 and lacks undefined components, such as the protein-free hybridoma medium. It can be used with a variety of different induction factors or cytokines to support differentiation of the ectoderm, mesoderm and endoderm lineages. The other serum-free medium used in this paper is the SFM prepared in house. It is a combination of the APEL formulation13 and the serum-free medium described by Bai et al.14 Usually, media prepared in house offers a more affordable option. The in vitro blood cell differentiation process is divided into key stages in which specific signals are provided for the mesodermal, hemangioblast/hemogenic-endothelium and hematopoietic stem/progenitor cell specification and further maturation. The initial stage of the hematopoietic differentiation was conducted by the embryoid body formation-based protocol. The formation of embryoid bodies was observed as early as 24 h after centrifugation (data not shown), as reported by Ng et al.,13 in which it was performed using the commercial differentiation medium STEMdiff APEL 2. When using the SFM prepared in-house, the formation of uniform embryoid bodies was not observed after 48 h, even with the addition of 0.05% polyvinyl alcohol to favor the formation of the embryoid bodies (Figure 2A). The differentiation media influenced the formation and uniformity of the embryoid bodies and this is probably related to the in-house preparation of the SFM medium. The variability between batches of human albumin and mainly, the preparation of the solution and the concentration of the PVA used, may have contributed to this. A combination of 0.5% human albumin and 0.05% PVA was used in the formulation of the SFM, as indicated by Ng et al.13 Authors have reported that the addition of albumin improves the growth and health of cells in culture, but the addition of albumin into the culture medium brings greater variability, which may compromise the reproducibility of differentiation experiments.15,16 This concentration of albumin and PVA is optimized for mesodermic induction, which is compromised when greater proportions of PVA are used. However, as shown in this paper, the less uniform embryoid bodies allowed for the emergence of supernatant and positive cells for hematopoietic markers.
Additionally, both media provided the emergence of CD34+ cells and can be used in hematopoietic differentiation (Figure 3). The percentage of CD34+ cell obtained with the iPSC_scd cell line on day 8 of differentiation were significant and corresponds to that reported in the literature, using ESC and iPSC from healthy donors and sickle cell anemia patients.10,14,17,18 Interestingly, only the differentiation with the SFM generated CD45+ cells. This hematopoietic marker was not detected when the cells were cultivated in the STEMdiff APEL 2 medium, but we did not rule out the possibility that the marker would be detected if the culture were prolonged. Throughout our study, double-positive CD34+/CD45+ populations of cells cultivated in the SFM were identified between days 8 and 14 of differentiation (Figure 4). Kennedy et al. (2012) detected CD45+ cells from day 13 of the ESC differentiation, when they found approximately 21.7% of positive cells that increased on day 15, reaching about 28.4% of CD45+ cells in embryoid bodies.19 Kobari et al. performed the differentiation of iPSC cells from the sickle cell anemia patient through the formation of embryoid bodies and detected, after 27 days of culture, approximately 65% of CD45+ cells and 3% of CD34+/CD45+ cells.9 Reis et al. (2017) obtained from embryoid bodies of the iPSC_scd cell line approximately 38% of CD34+/CD45+ cells on day 8 of the differentiation.10
The CD43 is one of the most prevalent transmembrane leukocyte sialoglycoproteins expressed in cells of the hematopoietic lineage, including hematopoietic stem cells. In this study, the iPSC_scd cell line differentiated in the SFM showed an increase in CD43+ cells from day 8–14 of the differentiation. This marker appeared one day earlier than Kennedy et al. had reported with the ESC embryoid bodies.19 A similar percentage of CD43+ cells was detected, as compared to what Reis et al. obtained from embryoid bodies of the iPSC_scd cell line with the STEMdiff APEL medium.10 Furthermore, we obtained CD43+/CD45+ cells, of which approximately 42.28 ± 36.88% (N = 3) of the cells were double-positive on day 14 (Figure 4). We also demonstrated the potential of these cells to form hematopoietic colonies in the methylcellulose culture. In our assays of colony formation with embryoid body cells collected on day 8 of differentiation with the SFM medium, it was possible to observe the formation of hematopoietic colonies, including colonies of the erythroid lineage (Figure 5A). Kessel et al., working with the iPSC from a healthy donor, determined that CD43+ cells are the population of cells that give rise to erythropoiesis in vitro.20 They also demonstrated, by means of a colony formation assay, that the fraction of hematopoietic stem cells/progenitors with multipotent potential is enriched in the CD43+/CD45+ cell population.
When cultivated in the STEMdiff APEL 2 medium, the iPSC_scd cell line did not show a significant increase of CD43+ cells throughout the differentiation (Figure 3). We observed the potential for colony formation in embryoid body cells with 8 days of hematopoietic development, when cultivated in the STEMdiff APEL 2 medium. However, with these cell populations, with a high percentage of CD34+ cells, low percentage of CD43+ cells and without the presence of the CD45 marker, no erythroid colonies were observed among the generated hematopoietic colonies (Figure 5B). The formation of hematopoietic colonies from day 8 embryoid bodies in the absence of CD45+ cells corroborates the findings of Wang et al. and Vodyanik, Thomson and Slukvin, in which the authors verified that the embryoid bodies analyzed between days 7 and 10 of development already contained competent hematopoietic precursor cells, even in the absence of the CD45 label expression.21,22 Vodyanik, Thomson and Slukvin demonstrated that hematopoietic progenitors derived from the ESC can be identified by the CD43 marker and that the selection of CD43+ separates the hematopoietic cells CD34+ from the endothelial cells CD34+/CD43−/KDR+/CD31+.22 Kennedy et al. analyzed the CD34/CD43 cell populations for the potential for colony formation.19 According to the authors, the CD34+/CD43− and CD34−/CD43− cells did not generate any colonies.
The second stage of the differentiation is the erythroid maturation. The embryoid bodies were cultivated in maturation medium, prepared with the SFM culture supplemented with cytokines for the induction of erythroid differentiation. The cells were matured in plates treated with gelatin to provide better hematopoietic development of progenitor cells. We observed cells adhered to the culture and the expansion of rounded and semi-adherent cells with morphology similar to hematopoietic stem cells/progenitors 6 days after the beginning of culture in the maturation medium (Figure 6), similarly to those that Xie et al. observed after 4 days in cultivation on the extracellular matrix.23 Other research groups demonstrated the generation of cell populations with erythroid morphology from ESC and iPSC patient-specific after 10–15 days in the maturation medium.12,24
In addition to the morphology, we analyzed the presence of CD45 and CD34 in the differentiated cells. Our results showed an increase of CD45+ cells in all experiments performed and a decrease in CD34+ cells after culture in the maturation medium (Table 1), suggesting the emergence of the proerythroblast population. The CD45 is expressed in megakaryocyte-erythroid progenitor cells and in the pro-erythroblast, and the CD34 expression is high in megakaryocyte-erythroid progenitor cell populations and low in the proerythroblast population. Pro-erythroblasts also show high expression of CD36, CD71 and CD235a, which are absent in megakaryocyte-erythroid progenitors.25 In the present study, cells cultured for 15 days in the maturation medium generated 48.28% of CD43+/CD45+ double-positive cells, with 90.02% of CD45+ cells, 40.33% of CD36+ and 66.17% of CD71+. However, no significant percentages of CD235a+ cells were detected at the time we performed the analysis. Kobari et al. (2012) performed the differentiation of iPSC cells with the sickle cell anemia mutation for a longer time, about 25 days in the erythroid differentiation medium, and detected a high percentage of positive cells for the three erythroid markers, including approximately 98% for CD235a+.9 Kessel et al. detected, at the end of 18 days of healthy donor iPSC in the differentiation medium for erythroid induction, almost all CD71+ and CD235a+ cells.20 These results suggest that, after 15 days of culture in the erythroid differentiation, the cells from iPSC_scd showed commitment to the erythroid lineage.
As evidenced in this study, it is possible to efficiently generate hematopoietic progenitor cells using a simple and inexpensive house-prepared medium. We used an iPSC derived from a sickle cell anemia donor to develop an efficient model of sickle cell anemia capable of generating hematopoietic progenitor cells and erythroid progenitors in vitro through the formation of embryoid bodies. Thus, we determined that the SFM differentiation medium prepared in house and supplemented with cytokines promoted the generation of the highest percentage of CD34+/CD45+ and CD43+/CD45+ cells in embryoid bodies cultivated for at least 8 days. These cells showed a potential for the formation of hematopoietic colonies, including erythroid colonies, when cultivated for a further 14 days in a semi-solid medium. The culture of the embryoid body cells for at least another 10 days in the erythroid maturation medium promoted the emergence of CD36+, CD71+, and CD235a+ populations.
ConclusionsIn summary, this report described the differentiation of sickle cell anemia patient-derived iPSC for the generation of hematopoietic stem/progenitor cells and erythroid progenitors, using a simple and inexpensive house-prepared medium. Our in vitro erythroid differentiation system provides a simple platform with a similar hematopoietic differentiation potential for reported human pluripotent stem cell systems. Furthermore, this platform could allow for disease modeling and the development of novel therapies for sickle cell disease.
Data availabilityThe data used to support the findings of this study are available from the corresponding author upon request.
Conflicts of interestThe authors declare that there is no conflict of interest regarding the publication of this manuscript.
The authors thank Patricia Viana, Camila Menezes and the Laboratory of Flow Cytometry for the help and discussions on the flow cytometry analysis. The authors also thank Andy Cumming for the English review. Financial support for this work was performed by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (process number 2016/08373-9), Centro de Terapia Celular (CTC) - FAPESP [process number 2013/08135-2], Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) [process number 141058/2016-1], Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and Blood Center of Ribeirão Preto, Brazil.
The following is Supplementary data to this article:
